|
UV GUIDE
UK
Advances
in Reptile Lighting
A
resource for all reptile keepers
|
|
The
Transmission of Ultraviolet Light
through
Reptile Skin Shed.
Simple
"kitchen experiments" to assess the relative amounts
of UVB light reaching the deeper layers of the skin in different
species of lizard and snake.
Reptile
Skin - The
Investigation - Results:
UVB readings - Spectrometer
recordings
Discussion
on results from different species
|
|
Introduction
When
studies were made on vitamin D synthesis in human skin,
it was soon discovered that the colour of skin you have
determines how sensitive it is to ultraviolet light. This
is because the brown pigment, melanin, which is found in
the skin, protects it from sun damage (eg. sunburn) by blocking
UV light. So, if you are dark-skinned, you are less likely
to get sunburnt; but your skin is not as efficient as that
of a fair-skinned human at producing vitamin D3 in sunlight.
If you are dark-skinned, and only receive a little UV light,
you are more likely than a fair-skinned person to suffer
from vitamin D3 deficiency.22
Now,
different species of reptiles have very different skin types.
Some sun-worshipping species, like bearded dragons and uromastyx,
have thick armour; some nocturnal geckos have delicate skin
like pink tissue paper. We wondered whether we could find
out how much UV is being absorbed by different types of
reptile skin.
This
has practical application. If, for example, a UVB lamp is
emitting 100uW/cm² at 12", and two lizards sit under
it at that distance, will they both be able to make the
same amount of vitamin D3? Obviously, the answer is no,
if one has skin which blocks more UVB than the other. To
get the required daily amount of vitamin D3, reptiles which
have less sensitive skin may well need either higher UVB
levels... or more time under the lamp.
We decided
to find out more about this, and first, we took a closer
look at reptile skin....
About
Reptile
Skin
 Reptile
skin, like that of many vertebrates, has two principal layers:
the dermis, which is the deeper
layer of connective tissue with a rich supply of blood vessels
and nerves, and the epidermis which
in reptiles consists of up to seven sub-layers or "strata"
of closely packed cells, forming the body's outer protective
coating. 46 Reptile
skin, like that of many vertebrates, has two principal layers:
the dermis, which is the deeper
layer of connective tissue with a rich supply of blood vessels
and nerves, and the epidermis which
in reptiles consists of up to seven sub-layers or "strata"
of closely packed cells, forming the body's outer protective
coating. 46
The
epidermis has no blood supply, but its innermost living
cells obtain their nourishment by the diffusion of substances
to and from the capillaries at the surface of the dermis
directly beneath them. It is on the cell membranes of these
cells that pre-vitamin D3 is synthesised from the cholesterol
precursor, under ultraviolet light. Once formed there, the
pre-D3 is ejected from the cell membrane to the intercellular
fluid where it is isomerised to vitamin D3. This diffuses
into the capillaries of the dermis, is picked up by the
D3 plasma binding protein and carried into the body.23
The
ultraviolet light reaching these cells has to come through
the outermost layers of the epidermis first. The epidermis
of different reptiles varies widely in its thickness and
pigmentation. (Melanin, responsible for brown colouration,
is found in the epidermal cells and absorbs UV light. In
reptiles the other colours are derived from cells in the
dermis beneath, which may contain pigments producing, for
example, reds and yellows; or reflect certain wavelengths
of light, giving, for example, iridescent blues).47
The
seven epidermal layers are
1. The
stratum germinativum, the deepest layer of living
cells which have the capacity for rapid cell division;
and
the six layers which form each "epidermal generation"
- the old and new skin layers - which are:
2 -
3. The clear layer and lacunar layer,
which mature in the old skin layer as the new skin is growing
beneath.
4 -
6. The alpha layer, the mesos layer
and the beta layer; these layers consist of cells
which are becoming keratinised, with production of two types
of keratin (alpha and beta keratin). These cells are thus
being transformed into a hard protective layer.
7. The
Oberhautchen layer, which forms the toughest outermost
layer of keratinised, dead skin cells.
In
most mammals, the structure of the epidermis is less complex
and the outermost, dead skin cells are constantly flaking
off (as "scurf"); this protective layer is constantly
replaced from below. The deepest layer of cells, the stratum
germinativum, is constantly dividing and multiplying, and
so all the layers are "on the move" outwards.
In reptiles,
however, this cell division in the stratum germinativum
only occurs periodically, and when it does, all the layers
above it, in the area where the cell division occurs, are
replaced in their entirety - the reptile literally grows
a second skin, underneath the old skin, and then "sheds"
the old one.
Some
lizards (such as the leopard gecko) and snakes, shed the
skin from the entire body all at once. Many other lizards
shed the skin in large patches; bearded dragons, for example,
may only shed the skin across the back, or the tail, or
the legs at any one time. (Figs. 2
and 3.)
About
2 weeks before the reptile sheds its skin, the cells in
the stratum germinativum begin active growth and a second
set of layers form slowly underneath the old ones. At the
end of this time the reptile effectively has a double skin,
and this can be seen as a darkening in colour, in many species.
Then,
the cells in the lowest layers of the old skin, (the clear
and lacunar layers) and the Oberhautchen layer of the new
skin below, undergo a final maturation and a so-called shedding
complex forms. Fluid is exuded and forms a thin liquid layer
between them. This gap between the two "skins"
gives the familiar milky appearance to a shedding reptile.
Enzymes in this fluid break down the connections between
the two layers; the old skin lifts; and the reptile actively
removes it. 46,48,49,50.
A
useful diagram which
shows the structure of the epidermis of a reptile, just
before the skin is shed, may be viewed from the University
of Michigan Museum of Zoology's The Animal Diversity
Web, by clicking on this link:- Reptile
epidermis (opens
in a new window)51.
(We cannot show it on this site for copyright reasons)
|
|
The
Investigation
Two
related studies by Carman et al 9
and Ferguson et al 16
compared the vitamin D3 production of the skins of four
species of lizard - a crepuscular house gecko, a shade-dwelling
anole, an anole which basks in sunshine, and the sun-dwelling
Texas spiny lizard. They found that the amount of vitamin
D3 production was inversely related to the amount of exposure
to UVB light the reptiles would normally receive in the
wild. The Texas spiny lizard skin produced the least; in
a high UVB environment, this thick skin might well be resistant
to UV damage and yet still produce sufficient vitamin D3.
The house gecko skin was the most sensitive; presumably
this gecko would be able to make the most of the smallest
amounts of ultraviolet light that came its way.
We wondered
whether the reason for the variation in sensitivity of the
skin to UV light was simply due to reduced UV penetration
of the skin in the reptiles which are regularly exposed
to higher levels of UV light. Casual observation suggests
that species of reptile which regularly expose themselves
to full sunshine, in hot dry environments (eg. uromastyx,
chuckwallas, bearded dragons) seem to have thicker,
tougher skins than crepuscular species from similar environments
(eg. leopard geckos) or diurnal reptiles which inhabit
shadier areas (eg. rainforest chameleons). No doubt,
the heavier body armour of the former species protects the
rest of the body from water loss and from intense solar
radiation. However, these species presumably have similar
vitamin D3 requirements to their cousins which live under
lower levels of light. To make the
required amount of vitamin D3 each day, would sun dwellers
actually need full sun, for sufficient UV to penetrate the
armour? And in the case of crepuscular reptiles, would the
skin allow even the smallest amounts of UV through?
To test
this theory fully would presumably require the sampling
of living skin removed from reptiles, and comparison of
the UV transmission through the cell layers. We reasoned
that although shed skin is not structurally identical to
unshed skin, and any comparison would therefore be crude
at best, there were sufficient similarities between the
two to make this very non-invasive
skin sampling technique worthwhile.
 We
have therefore begun collecting fresh
shed skin samples from a range of lizard species.
When the shed skin first lifts, and is pulled off by the
shedding lizard, it is still hydrated, soft and flexible.
If pieces of this are rescued before they dry out or become
damaged, they can be used to gain a very crude approximation
of the UVB transmission of that reptile's skin. We
have therefore begun collecting fresh
shed skin samples from a range of lizard species.
When the shed skin first lifts, and is pulled off by the
shedding lizard, it is still hydrated, soft and flexible.
If pieces of this are rescued before they dry out or become
damaged, they can be used to gain a very crude approximation
of the UVB transmission of that reptile's skin.
All
that is required is to place the sample between a UVB meter
and a known UVB source, and record the observed reduction
in UVB output.
By placing
the skin sample between the sensor of a spectrometer and
a known light source, it is also possible to observe the
absorption of light (across all wavelengths) by the piece
of skin.
|
|
Method
(1)
UVB Meter Readings
A Solarmeter
6.2 meter, recording UVB output in uW/cm², was used for
all tests. The meter was fixed in position below various
known sources of UV light, including the sun and a range
of UVB fluorescent tubes and mercury vapour lamps from the
2005 Survey. Readings were taken before and after the fresh
shed skin samples were placed directly over the sensor of
the meter in such a way that they acted as a filter for
the UVB light. Care was taken to ensure undamaged parts
of the sample completely covered the sensor on each occasion.
One
set of recordings are shown in Fig.5.
To the left is the unobstructed Solarmeter under the UV
lamp, at a distance at which the output is 100uW/cm². In
the centre, a sample of shed skin from a leopard gecko (Eublepharis
macularius) is over the sensor, and a transmission of
38% is recorded. To the right, this has been replaced by
a sample of bearded dragon (Pogona vitticeps) shed
skin. Transmission is now only 2%.

A further
set of readings, for a small number of samples, was taken
using a Solarmeter 6.4, which has a narrower band of sensitivity
and records UVB output in terms of International Units of
D3 per minute (as produced by "typical" human
skin - see the manufacturer's details for an explanation
of this). (http://www.solarmeter.com/model64.html)
(2)
Spectrometer Recordings
The
spectrometer used for all tests was an Ocean Optics Inc.
USB 2000 Fibre Optic Spectrometer. (http://www.oceanoptics.com)
The light source used was a ZooMed Reptisun 10.0 fluorescent
tube with known output (ref. BZ9). The spectrometer sensor
was clamped in place at a distance of 10cm from the light
source and recordings taken before and after the skin samples
were placed immediately in front of the sensor.
|
|
Results
(1)
UVB Meter Readings
Tables
1a
(lizard skin) and 1b
(snake skin) show the results obtained from the
Solarmeter
6.2 meter, when sampling skin shed from
more than 60 animals from 23 different species. Where more
than one sample was tested within a species / group, the
number of samples is indicated in brackets and the average
of the results is given. We are grateful to members of UVB_Meter_Owners
group who have expanded our original study, contributing
samples and readings for this section. (Please see Acknowledgements
at foot of page)
| Table
1a : lizards |
| Species
/ group |
Origin
of skin shed sample |
Percentage
transmission |
| Green
iguana - adult male |
tail
(underside) |
0% |
| Green
iguana - adult male |
tail
(upper black banded area) |
1% |
| Bearded
dragon - adults |
back
(3) |
4% |
| Bearded
dragon - adult |
|
5% |
| Green
iguana - adult male |
tail
(upper green banded area) |
5% |
| Beaded
Lizard |
belly |
5% |
| Argentinian
black-and-white tegu |
back |
8% |
| Green
iguana - two young adults with MBD |
chest
(2) |
9% |
| Gila
Monster |
|
10% |
| Ornate
uromastyx - young adult male |
back
(dark markings) |
11% |
| Argentinian
black-and-white tegu |
side
/ belly |
12% |
| Bearded
dragon - adult |
belly |
15% |
| Ornate
uromastyx - young adult male |
back
(light markings) |
16% |
| Bearded
dragon - adult |
under
chin |
16% |
| Parson's
chameleon |
belly |
16% |
| Panther
chameleon - old female |
side |
19% |
| Panther
chameleon - male |
side
(3) |
20% |
| Parson's
chameleon |
tail
(2) |
20% |
| Globifer
chameleon |
side |
21% |
| Yemen
(Veiled) Chameleon - female |
side |
22% |
| Parson's
chameleon |
side |
24% |
| Oustalet
chameleon |
side
(3) |
26% |
| Green
Basilisk |
|
26% |
| Leopard
gecko |
head |
33% |
| Panther
chameleon - gravid female |
side |
34% |
| Crested
gecko |
head
/ back (2) |
35% |
| Leopard
gecko |
(2) |
37% |
| Leopard
gecko |
back
(3) |
39% |
| Leopard
gecko |
tail
-upper surface (2) |
39% |
| Leopard
gecko |
axilla
/ belly (2) |
41% |
| Panther
chameleon - subadult female |
side
(2) |
41% |
| Panther
chameleon - subadult male |
side |
42% |
| Panther
chameleon - few weeks old |
side
(2) |
51% |
| Table
1b : snakes |
| Species
/ group |
Origin
of skin shed sample |
Percentage
transmission |
| Sarong
Green Tree Python (dusty sample) |
|
13% |
| Argentine
Boa - 1 male, 1 female |
(2) |
18% |
|
Ball Python - normal pigmentation |
|
27% |
| Dumeril's
Boa |
|
29% |
| Ball
Python - Piebald |
pigmented
area |
32% |
| Garter
snake |
back |
38% |
| Ball
Python - ghost |
|
45% |
| Grey-Banded
Kingsnake |
|
45% |
| Grey-banded
Kingsnake |
back |
48% |
| Albino
Nelson's Milksnake |
|
48% |
|
Ball Python - albino |
|
48% |
| Sinaloan
Milksnake |
|
49% |
| Ball
Python - Piebald |
white
area |
53% |
| Variable
(50/50 Californian) Kingsnake
|
back |
54% |
| Garter
snake |
belly |
56% |
Table
2 shows the results obtained using a Solarmeter
6.4 meter to compare the transmission of UVB
light in the wavelengths which are optimal in producing
vitamin D3. (290 - 315nm).
| Species |
Origin
of skin shed sample |
Percentage
transmission |
| Bearded
dragon |
back |
4% |
| Leopard
gecko |
head |
27% |
| Leopard
gecko |
back
(2) |
35% |
| Leopard
gecko |
tail
(2) |
34% |
| Leopard
gecko |
axilla
/ belly (2) |
39% |
(2)
Spectrometer Recordings
The
spectrometer is able to process the data in several ways.
Firstly we acquired a simple set of spectrograms which show
the spectral power distribution of the lamp with and without
the skin samples in place. Although the spectrometer provides
data across all wavelengths from UVC to infra-red, Fig.6
shows the results in the UVB wavelengths only.
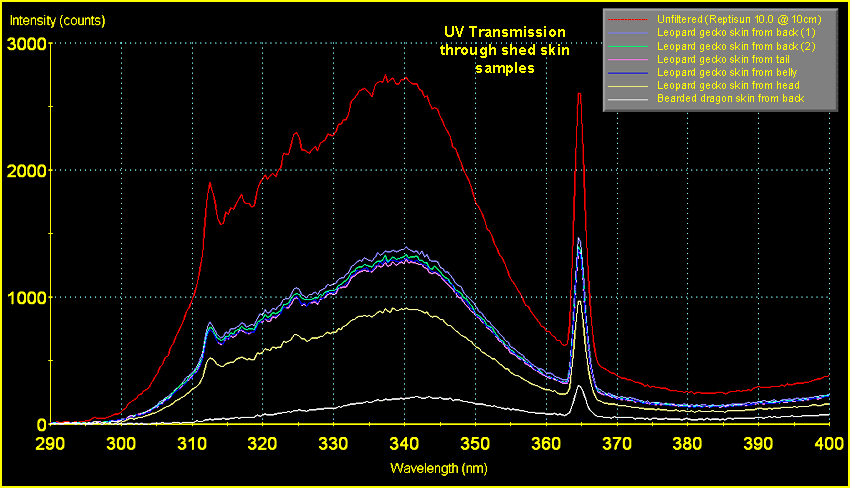
The
scale of this graph does not permit close inspection of
the data in the range of wavelengths responsible for D3
synthesis (290 - 315nm; peak production being at 297nm).
This can be seen a little more clearly in Fig.
7, below.
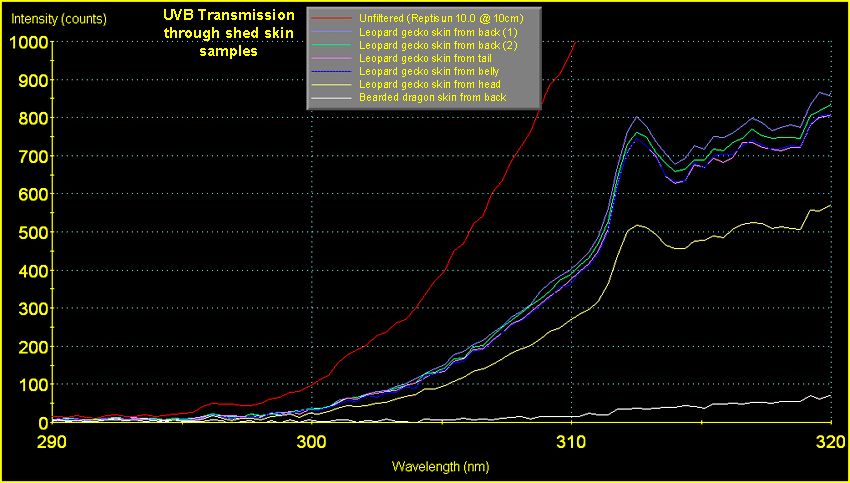
|
|
Discussion
The
results clearly demonstrate a huge variation between species,
in terms of the transmission of UV light through shed skin.
Although we have no proof that the UV transmission in any
given species is directly related to the levels of UVB in
the natural environment of each species, it is interesting
to ponder the fact that there does appear to be a surprising
correlation...
Bearded
dragons (Pogona vitticeps),
a species representing the "sun worshippers",
live in arid Australian scrubland and bask in the open sun.
The shed skin from their backs blocks all but 3-6% of the
total UVB according to the Solarmeter 6.2; these figures
are confirmed by the spectrograms which suggest an even
greater absorption of UVB at the wavelengths which are responsible
for D3 synthesis. Presumably, they receive enough high-UV
direct sunlight every day to make sufficient vitamin D3
from just the small percentage of it which reaches the deeper
cell layers of the epidermis. Skin from their undersurfaces
(belly and throat) seems less protective; the samples allowed
up to 16% total UVB through; we might speculate that these
areas will receive less direct sunlight and so this optimises
the D3 production from these more sheltered parts of the
body.
In captivity,
we might expect this species to have a very high UVB requirement,
since so little of what is provided actually penetrates
the skin.
 Green
iguanas (Iguana iguana),
live in tropical rainforest, where they normally bask for
long periods in full sunlight early and late in the day.
During the heat of the day, they move in and out of leafy
shade; here they are exposed to reflected and diffused UVB
in similar amounts to that of the morning and evening sun
in which they bask 27,32.
The shed
skin from the tail of an iguana is strongly banded, matching
the black melanin bands in the dermis. (Melanophores in
the dermis produce cellular extensions which infiltrate
the epidermal layers; these leave small amounts of melanin
in the shed skin.11) The
shed skin from a mature adult iguana (which had lived in
a naturalistic, high UVB environment for years) was very
thick and heavily keratinised; even the unpigmented areas
on the dorsal surface blocked all but 4-6% of the total
UVB; areas with melanin only transmitted 0-1%. Shed skin
from the chest area of two young adults under treatment
for metabolic bone disorder (MBD) was slightly more permeable
to UV light, transmitting 7% and 11% respectively. Whether
their age, or history of inadequate exposure to UV light
has any bearing upon the result is not known. Even so, 7%-11%
transmission is low. In captivity, this species might, like
the bearded dragon, be expected to have a very high UVB
requirement, since such a small proportion penetrates the
iguana's armoured hide. Green
iguanas (Iguana iguana),
live in tropical rainforest, where they normally bask for
long periods in full sunlight early and late in the day.
During the heat of the day, they move in and out of leafy
shade; here they are exposed to reflected and diffused UVB
in similar amounts to that of the morning and evening sun
in which they bask 27,32.
The shed
skin from the tail of an iguana is strongly banded, matching
the black melanin bands in the dermis. (Melanophores in
the dermis produce cellular extensions which infiltrate
the epidermal layers; these leave small amounts of melanin
in the shed skin.11) The
shed skin from a mature adult iguana (which had lived in
a naturalistic, high UVB environment for years) was very
thick and heavily keratinised; even the unpigmented areas
on the dorsal surface blocked all but 4-6% of the total
UVB; areas with melanin only transmitted 0-1%. Shed skin
from the chest area of two young adults under treatment
for metabolic bone disorder (MBD) was slightly more permeable
to UV light, transmitting 7% and 11% respectively. Whether
their age, or history of inadequate exposure to UV light
has any bearing upon the result is not known. Even so, 7%-11%
transmission is low. In captivity, this species might, like
the bearded dragon, be expected to have a very high UVB
requirement, since such a small proportion penetrates the
iguana's armoured hide.
 The
Argentinian Black-and-white Tegu (Tupinambis merianae),
a diurnal basking lizard, appears to have skin which protects
the lizard from most of the sun's UVB, somewhat similar
to that of the bearded dragon. The skin on the back is slightly
more transparent to UVB than bearded dragon skin, the skin
on the flanks and belly slightly less so. It is tempting
to speculate that this is the result of the wild tegu's
less harsh solar environment; they live in a temperate climate
in rough grassland and mountain valleys. The
Argentinian Black-and-white Tegu (Tupinambis merianae),
a diurnal basking lizard, appears to have skin which protects
the lizard from most of the sun's UVB, somewhat similar
to that of the bearded dragon. The skin on the back is slightly
more transparent to UVB than bearded dragon skin, the skin
on the flanks and belly slightly less so. It is tempting
to speculate that this is the result of the wild tegu's
less harsh solar environment; they live in a temperate climate
in rough grassland and mountain valleys.
 Ornate
Uromastyx (Uromastyx ornata) are
lizards which live in extreme desert conditions in their
native Egypt, Sinai and northern Arabia, in steep, rocky
ravines that are dry for most of the year. Unlike the previously-mentioned
species (which apparently spend much of their time in the
daylight) ornate uromastyx, when not basking or feeding,
shelter from the extremes of the desert environment in crevices
in rocks. Other uromastyx species inhabit burrows which
they dig in the ground.53
The sample of shed skin tested was more transparent to UVB
than bearded dragon skin but whether this is related to
the difference in behaviour is not known. In captivity,
they might be expected to have a high requirement for UVB
light - but also the need to be able to shelter from it
in hides or burrows, as they would in the wild at certain
times during the day. Ornate
Uromastyx (Uromastyx ornata) are
lizards which live in extreme desert conditions in their
native Egypt, Sinai and northern Arabia, in steep, rocky
ravines that are dry for most of the year. Unlike the previously-mentioned
species (which apparently spend much of their time in the
daylight) ornate uromastyx, when not basking or feeding,
shelter from the extremes of the desert environment in crevices
in rocks. Other uromastyx species inhabit burrows which
they dig in the ground.53
The sample of shed skin tested was more transparent to UVB
than bearded dragon skin but whether this is related to
the difference in behaviour is not known. In captivity,
they might be expected to have a high requirement for UVB
light - but also the need to be able to shelter from it
in hides or burrows, as they would in the wild at certain
times during the day.
Pigmented
areas of the shed skin, containing melanin, blocked UVB
more strongly than unpigmented areas in a similar way to
that seen with iguana skin.
 Leopard
Geckos (Eublepharis macularius)
are crepuscular although they have often been observed basking
in captivity, and it is also likely that small amounts of
UV light penetrate their daytime shelters in the wild. All
the samples taken from the backs of four different geckos
proved remarkably similar, transmitting 37 - 44% total UVB
(measured with the Solarmeter 6.2, confirmed by the spectrograms)
and 34-35% in the D3 range (measured with the Solarmeter
6.4, also confirmed by the spectrograms). The skin shed
from a leopard gecko's back transmits up to 14 times more
UVB than the shed from a bearded dragon's back. It seems
likely that this skin would respond efficiently to low levels
of UV light, since even small amounts of light would reach
the deeper layers where D3 synthesis can take place. Leopard
Geckos (Eublepharis macularius)
are crepuscular although they have often been observed basking
in captivity, and it is also likely that small amounts of
UV light penetrate their daytime shelters in the wild. All
the samples taken from the backs of four different geckos
proved remarkably similar, transmitting 37 - 44% total UVB
(measured with the Solarmeter 6.2, confirmed by the spectrograms)
and 34-35% in the D3 range (measured with the Solarmeter
6.4, also confirmed by the spectrograms). The skin shed
from a leopard gecko's back transmits up to 14 times more
UVB than the shed from a bearded dragon's back. It seems
likely that this skin would respond efficiently to low levels
of UV light, since even small amounts of light would reach
the deeper layers where D3 synthesis can take place.
The
skin shed sample from the head of the leopard gecko was
less permeable to UV light. It looks darker to the naked
eye, as well, and the pigmented spots are more clearly seen.
Why the skin of the head should provide better UV shielding
than skin on the rest of the body is unknown. Although skin
from the head was only sampled from one gecko, it is interesting
to note that the skin from the back of this animal was the
most transparent to UV of all the gecko skin tested, making
the contrast with the head more remarkable.
In captivity,
we might expect this species to benefit from low levels
of UVB, but high levels might be harmful, since such a large
proportion of it penetrates the skin.
Chameleons
appear to fall between the two extremes.
 Panther
Chameleons (Furcifer pardalis) and Oustalet’s Chameleons
(Furcifer oustaleti)
are found in open forest as well as trees and bushes alongside
Madagascan roads. Both species are often found in the same
trees. Panther
Chameleons (Furcifer pardalis) and Oustalet’s Chameleons
(Furcifer oustaleti)
are found in open forest as well as trees and bushes alongside
Madagascan roads. Both species are often found in the same
trees.
Their environment is bright with high levels of UV, however
neither species basks to the extent that a bearded dragon
does. In addition they are arboreal, and as vegetation absorbs
UVB, parts of their day are spent within trees and bushes
in sheltered UV areas.
Oustalet’s
are larger than panthers with thicker, tougher skin. Surprisingly,
testing various skin samples for males of both species using
the Solarmeter 6.2 showed Oustalet’s skin has a higher UVB
transmission rate. This would suggest the Oustalet’s chameleon
may have a higher UVB requirement than the panther chameleon.
Adult
Panther Chameleons showed
profound differences depending upon their sex and breeding
condition. A gravid female had a transmission rate of 34%;
an old, non-breeding female and an adult male both had skin
transmitting only 19%. It is possible the gravid female's
increased requirement for vitamin D3 and calcium is, in
some unexplained way, altering the skin's absorption of
UVB.
Juvenile
Panther Chameleons
Skin from several baby panthers were tested and UVB transmission
was found to be around 50%, and a sample from a sub-adult
male transmitted 42%. These readings are significantly higher
than for the adults tested. A number of reasons for this
difference are suggested below:
 Body
size / overall skin thickness. Body
size / overall skin thickness.
A panther’s skin is thinner when a baby than as an adult.
However when comparing skin thickness and UV transmission
across species, both Oustalet’s and Parson's skin are
thicker than panthers and yet both allow more UV through.
Therefore it seems likely this extra transmission is for
reasons other than, or at least not limited to, body size.-
Requirements for Growth.
Baby chameleons are fast growing and the demands of laying
enough calcium down for strong bones means their D3 requirements
and therefore their UVB requirements are higher than those
of adults.
- Basking
Preferences.
At times of higher D3 requirements (e.g. gravid female)
adults increase the time they spend basking, and the resulting
additional UVB exposure facilitates higher vitamin D3
photobiosynthesis. Unfortunately basking in the wild carries
a cost, namely, exposure to predators. (It’s also worth
mentioning that basking reptiles tend to bask longer and
have slightly higher body temperatures in captivity than
in the wild, possibly because the threat of predation
is removed.)
The dangers of predation for a baby chameleon would be
higher than for an adult. In fact it would seem so much
more dangerous that baby chameleons inhabit a different
area from the adults. The wild Jackson’s chameleons from
the back garden of Mary Lovin in Hawaii 52
illustrate this point well. Adult Jackson’s tend to inhabit
the middle to top of the trees and bushes. The babies
spend their time closer to the ground in more sheltered
low-level bushes.
The babies by habit and habitat are not exposed to the
same levels of direct sunlight as the adults. It seems
possible that the higher UVB transmission in the babies’
skin would compensate for the more sheltered environment,
where there is less UVB light, and also for the babies’
higher demand for vitamin D3.
In captivity, we should consider the extra UVB requirements
that a baby may have compared to the adult. However, sufficient
UV shelter (generally provided by live plants) and a UV
gradient must be provided to allow for proper self-regulation
of exposure.
Parson’s
Chameleons (Calumma parsonii parsonii) and Globifer
Chameleons (Calumma
globifer) are two closely related species
inhabiting Madagascan rainforests well within the canopy.
They are shade dwelling, slow growing chameleons, which
are not considered basking animals.
Skin samples tested allowed moderate levels of UVB through,
with readings close to the Oustalet’s. But because Parson's
and Globifers come from a lower UVB environment and do not
bask (at least to any great extent), their UVB requirements
in captivity should not be considered similar to Oustalet’s.
In the wild they would be receiving fairly even ambient
levels of UVB all day, rather than moving in and out of
the light, as a basking species would do.
Yemen
or Veiled Chameleons (Chamaeleo calyptratus)
are native to Yemen and southern Saudi Arabia, and are found
in a wide range of habitats including dry plateaus, mountains,
and river valleys. They are diurnal, usually found in bushes
and shrubs in the daytime, retreating into shade during
the middle of the day. Skin samples allowed moderate amounts
of UVB through. These chameleons live in an environment
with high levels of UVB in the sunlight, but because they
are predominently shade dwellers their UVB requirements
in captivity may be considered similar to those of Parson's
and Globifer chameleons.
Unlike the bearded dragons tested, chameleon skin gives
more uniform results, regardless of where on the body the
samples come from. This is probably because, being an arboreal
and lateral basker, the chameleon’s whole body would be
exposed to UVB.
Crested
geckos (Rhacodactylus ciliatus) originate
from New Caledonia where they can be found on the southern
half of the main island Grande Terre as well as on the neighbouring
islands of the Isle of Pines and Isle of Komoto.
Their natural habitat consists mainly of humid, lowland
forests, where they spend their days hiding in leaves on
trees, bushes and the forest floor. As arboreal geckos,
they can easily situate themselves in positions of direct
sunlight if required. Crested geckos have been observed
outdoors moving throughout the day, keeping their body exposed
to rays of natural sunlight.
  They
change colour at times, when moved from indoors to an outdoor
position in full sun and temperatures within their normal
range. Although the exact purpose or cause of colour change
is at present unknown, there are theories linking levels
of light with changes in skin colouration. The shades and
degree of colour can be very marked; one day changing to
much darker or vivid colours, but the next day changing
to very pale, almost off-white /beige even in the same sunlight
conditions. They
change colour at times, when moved from indoors to an outdoor
position in full sun and temperatures within their normal
range. Although the exact purpose or cause of colour change
is at present unknown, there are theories linking levels
of light with changes in skin colouration. The shades and
degree of colour can be very marked; one day changing to
much darker or vivid colours, but the next day changing
to very pale, almost off-white /beige even in the same sunlight
conditions.
In captivity, crested geckos can be observed following their
usual nocturnal behaviour, usually selecting a favourite
sleeping spot, within leaves. Out of a group of 2 males
and 3 females, over the winter period (with slightly lower
temperatures and a break from breeding) only one female
slept partially exposed to UV lighting. When temperatures
started to rise and mating began, all 3 females were observed
to sleep at least part of the day partially or fully exposed
to the UV lighting. In the previous summer, only 1 male
chose not to expose itself to the UV lighting on a daily
basis, within its vivarium. But when taken outside on a
sunny day, this gecko relished climbing to the top of its
tree and basking in full sunlight. All the group are proven
breeders.
Snakes
More research is needed as to the relationship between UV
light and vitamin D3 metabolism in the snake. Traditionally,
nocturnal and crepuscular snakes are not thought to require
UV light. However, there would not seem to be any reason
why they should not be able to utilise it for vitamin D3
synthesis. Is there any difference in transmission of UV
between the skin of nocturnal and diurnal species of snake?
Snakes
such as the forest-dwelling kingsnakes, which live in shaded
habitats and are often active at night, might be expected
to have skin which allows a high degree of UVB penetration;
and indeed, this does seem to be the case; the skin from
several species transmitted 45% or more of the UVB.
However,
the
inverse relationship between skin permeability and the amount
of sun to which the reptile exposes itself does not seem
as straightforward in snakes as it does in lizards.
Garter
snakes (Thamnophis sirtalis), for example, are diurnal
and often seen basking in morning sun, in a wide range of
habitats across Canada and the USA from Alaska to Texas.
Surprisingly, skin from the back of the snake would appear
to transmit a similar proportion of UV light as that of
the back of a leopard gecko. The skin on the belly was even
more translucent, allowing 56% of the UVB through.
The
skin of boas and pythons (with normal pigmentation) is much
more protective. The skin of the Argentine Boas (Boa
constrictor occidentalis), for example, permitted only
18% transmission. This is a nocturnal snake of the forest,
grassland and scrub of Paraguay and Argentina; one might
have expected a more "transparent" skin. It is
tempting to speculate that these snakes, although active
at night, do not hide away during daylight but rather, rest
in well-lit tree branches where they are exposed to significant
amounts of scattered UV light, if not direct sunlight.
Not
surprisingly, the skin of 'albino' animals, and patches
of unpigmented skin in piebald morphs, permitted a high
percentage of UVB penetration compared to normal, pigmented
skin. Such animals might be expected to be abnormally sensitive
to UV light; melanin is one of the skin's main defences
against UV light.
|
|
Further
food for thought/research needed
-
It would be interesting to be able to compare shed skin
from wild reptiles, exposed to natural sunlight, with
the shed skin from captive ones housed indoors with access
to varying amounts of UV light.
- “Tanning”
is a well-documented response in humans to an increase
in UV exposure. Does a similar process occur in reptile
skin?
- Do
animals that have been bred in captivity for many generations
have more transparent skin than their wild relatives?
-
Is there any difference in UVB transmission between sexes,
in species other than the Panther chameleon? Generally
females have a higher UVB requirement when gravid. Does
their skin transmit more UVB; or do they merely increase
the time spent basking?
-
Chameleons flatten their body when basking to create a
larger skin area, which traps heat quickly. This also
creates a wide skin surface for D3 photobiosynthesis.
The same chameleon walking around has a more tubular shape
and therefore a smaller surface area exposed to the sunlight.
Does this change in surface area have a direct effect
on D3 photobiosynthesis?
|
|
Acknowledgements
We are
grateful to the following members of the UVB_Meter_Owners
Group for submitting the additional samples and test results
which appear above:
Green
iguanas (Iguana iguana) - Robert MacCargar
Ornate
uromastyx (Uromastyx ornata) - Torey Lehman
Crested
gecko (Rhacodactylus ciliatus) - Angi
Grey-banded
kingsnake (Lampropeltis alterna) - Brian Stearns
Variable
(50/50 Californian) kingsnake (Lampropeltis getula x)
- Brian Stearns
Green
Basilisk (Basiliscus plumifrons); Gila Monster (Heloderma
suspectum); Beaded Lizard (Heloderma horridum);
Sinaloan Milksnake (Lampropeltis triangulum sinaloae);
Albino Nelson's Milksnake (Lampropeltis triangulum nelsoni);
Grey-banded kingsnake (Lampropeltis alterna); Ball
Pythons (Python regius); Dumeril's Boa (Boa dumerili);
Sarong Green Tree Python (Morelia viridis); Argentine
Boa (Boa constrictor occidentalis) - Jon Coote
We welcome
further contributions from members of the group.
|
|
© 2006 UVGuide.co.uk
|



















